| Shoot apical meristem
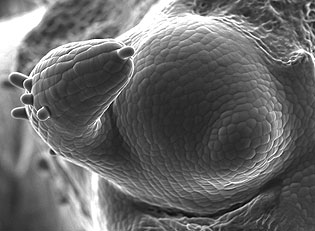
The vegetative shoot apical meristem consists of a small group of dividing cells, which give rise to leaf primordia in very regular and predictable temporal and spatial patterns.
Organ positioning, or phyllotaxis, is characterized by the divergence angles between the organs, the most common angle being 137.5°, the golden angle. We are interested in understanding the biochemical, physical and mathematical basis of this regularity. |
 |
 |
|