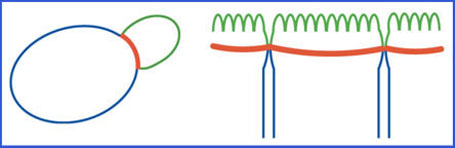
| Establishment of membrane fences and local cell wall deposition in plants In the simplest description of a eukaryotic cell, nucleus, cytoplasm and organelles are embedded in a phospholipidic bilayer, the plasma membrane. Proteins of different nature populate the plasma membrane and perform specific functions. Generally speaking, these populations of proteins are present all along the plasma membrane. However, some protein activities need to be delimited in space. For example, receptors present on the plasma membrane of a neuron must be localized to either the dendritic or the axonal end; in an epithelial cell, different transporters sit on either basolateral or apical membrane. A protein exclusively present in a subregion of the plasma membrane is said polarly – asymmetrically – localized. Molecular fences at the plasma membrane help maintain polarity by preventing free protein diffusion. Such molecular fences have been described in several cellular contexts: for example, diffusion between membrane compartments is prevented in budding yeast at the level of the bud neck and in epithelial cells, between apical and basolateral membrane.
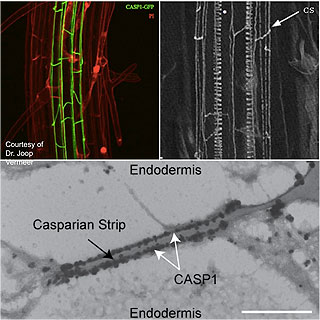
In plants, the only described example of membrane fence is found in the root endodermis, between inner and lateral plasma membranes. Recently, we have identified in A. thaliana a family of membrane proteins that define the endodermal membrane fence: CASPARIAN STRIP MEMBRANE DOMAIN PROTEINS (CASP1-5) localize at the root-shoot equatorial plane, splitting de facto the plasma membrane in two lateral domains. Besides establishing a fence at the plasma membrane, CASPs drive a local modification of the cell wall, the Casparian strips. CASP membrane domain and Casparian strips are perfectly aligned, and form adjacent 3D networks.
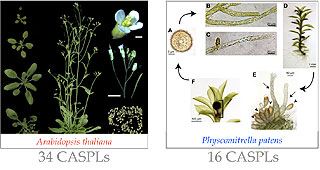
Are CASP-like as well able to make membrane fences? Are they involved in local cell wall modification? Can we use them to trace additional examples of membrane fences in plants? My lab is investigating the existence of membrane fences in plants by studying the localization of the CASPL proteins in a model dicotyledon, A. thaliana, and a model bryophyte, Physcomitrella patens. The ultimate scope of this characterization is the understanding of the molecular mechanisms that 1) generate membrane fences in plants (how do proteins localize in such structures? How are they maintained there?); 2) direct spatially restricted cell wall modifications. |
| Team |
Daniele Roppolo (Project leader) I have joined the IPS as independent project leader in February 2013. The Swiss National Science foundation (SNSF) funds our work through an AMBZIONE grant. I have an education as cell and molecular biologist, and I’m interested both in gene regulation and cell biology. I have acquired most of my competence in cell biology during my postdoc in Niko Geldner lab, University of Lausanne. My main contribution during this period was the identification of a family of proteins, the CASPs, that form a plasma membrane domain in the root endodermis; moreover, CASPs direct a local cell wall modification (Roppolo et al., 2011). During my PhD in Ivan Rodriguez lab, I worked on the understanding of the regulation of the V1r genes in the mouse. V1rs code for Vomeronasal pheromone receptors, which are important for several primary behaviours such as mating, sex discriminations, mother-newborn bonding. I showed that V1rs regulation leads to the transcription of a single allele from a single gene (out of 150) in a given olfactory neuron, and that the organization of V1rs in gene clusters helps ensure monogenic and monoallelic expression (Roppolo et al., 2007). |
Jacqueline Gheyselinck (Technician) I have worked as technician in several fields, from haematology to plant biology, and acquired international experience. In Ueli Grossniklaus lab, University of Zurich, I extensively used RNA in situ hybridization to unravel gene expression in ovules and embryos. At the University of Lausanne, I worked in the group of Uta Paszkowski, where I was responsible for the generation of transgenic rice plants. I have joined the IPS in March 2013; I’m working with Daniele to characterize CASPL expression patterns and cell wall modifications in caspl mutants. |